Table of Contents
Graphic mind interface
Goals:
Final goal : Graphic interface to mind based on subconsciousnes generated visual data showed in visual field ( mind eye ).
Experience
Experience over the last two years have shown many risks,side effects and lot of ways how not to do it.
Intention make this interface cause many poroblems its create intention/suggestions working in the subconscious which i long time not knew about.This suggestions cuase lowering of preception priority of physical senses and lowering significance of physical reality as a whole. These changes did not create the required interface (for reasons which i found out much later)but create interesting side effects. Lowering priority of physical reality freed resources for this perception used , this free resources began to be used subconsciousnes processes. Physical manifestations were more fatique and impaired senses especially vision. Mental symptoms increased perception of emotions, thoughts and emotions foreign to me, preception of “ghosts”, comunication with “ghosts” and different kinds of aspects of personality, better comunication with awareness. Especially in the beginning it was very difficult i had headaches and emotions with limited control over them.Gradually I learned how to navigate and working with them sensations became intense but controlable.I knew that essentially exhibit classic symptoms of schizophrenia but i began have state sufficiently under control to allow me to explore this state and its mechanisms deeper (dangerous but exciting journey). I have gained an interesting new perspective on how schizophrenia works which included a much broader view of different interactions at different levels. For example, as trigger “schizophrenia” can serve any phenomenon that reduces priority physicals or physiacal reality importance or something witch increse priority of nonplysical senses and reality.This lowering can by cased for exmple by depresion , deterioration of the physical senses , intention/suggestions. Recently, trying to fix the setting of priorities of physical senses and at the same time you maitain advantages nonphysical perception give me.
Notes
Hypangogic state
- 12-14Hz EEG spindles during the SO transition , but may reflect loss of consciousness of the external environment.
- Adiminution in the N3 ERP component may also index an attentional shift from external sensation tointernal imagery
- Occipital alphablocking with low-voltage, 4-7 Hz theta waves has also beenreported that appears to be associated with high rates of imagery recall
- EEG-derived dipoles indicate that spontaneous thoughts at SO differin localization (anterior vs. posterior) and frequency band(delta/theta and alpha primarily) depending upon the specificcontent under investigation (e.g., abstract vs. visual)
- Dream-like experiencesoccur most frequently in SO-stage 4 (flattening of the EEG to less than 20µV)

- frequency of kinesthetic images peaks in SO-stage 1 thendecreases steadily in later stages, whereas the opposite is truefor visual images
- Distribution of Modalities OverSO Stages
Of the 120 images, 80.9% (97) were classified as occurringin SO-stage 4 (EEG flattening) whereas 12.5% (15) wereclassified as occurring in SO-stage 5 (theta ripples). Onlyseven images (5.8%) occurred in SO-stages 1 or 2, a singleimage (0.8%) occurred in SO-stage 7, and no images occurredin SO-stages 8 and 9. * Spectral Powerof Kinesthetic and Visual Images
- 1) awake EEG samples, forbfrequencies faster than delta, had higher power than did corresponding image samples–regardless of whether thesewere kinesthetic or visual samples;
- 2) image samples had higher delta power than awake samples;
*indicated reduced power during image samples regardless of whether these were visual or kinesthetic. The effect was by farmost generalized for alpha, for which all 19 electrodessignificantly discriminated awake from imagery samples. *Findings from two previous self-observational studies(Nielsen, 1991, 1995) suggested that images produced withthis method are temporally-linked to the initiation of muscleatonia and/or phasic neuromuscular events such as limbtwitches and head jerks. If this is the case, then it is also likelythat atonia-linked movements experienced by our subjectsduring the task awakened them before they progressed beyondSO-stage 4 or, at most, 5.
Ganzfeld
Ganzfeld-induced hallucinatory experience, its phenomenology and cerebral electrophysiology
http://www.efectoganzfeld.com/uploads/5/3/0/3/5303662/ganzfeld.pdf
- The term ganzfeld originally denoted a homogeneous visual
field. By analogy, unstructured or de-structured stimulation can be applied to other sensory systems, e.g., auditory or tactile. Studies aiming at induction of ASC have been using ‘multi-modal ganzfeld’ (MMGF), i.e., simultaneous exposure to unstructured visual and auditory input.
- Further research focused mainly on the conditions of figure-
ground differentiation in the ganzfeld and colour perception (see Avant, 1965, for a review, cf. also Tsuji et al., 2004).
- In our earlier experimental studies (Wackermann et al.,
2002; Pu¨ tz et al., 2006) a red-coloured incandescent 60-W lamp, placed at a distancew120 cm from the eye-shields, was used as the light source; in recent studies, where a precise control of the ganzfeld colour is important, a computerdriven, xenon lamp based D-ILA projector has been used (Pu¨ tz and Wackermann, 2007). The choice of red colour reportedly (Cohen, 1958) facilitates the observers’ ‘immersion’ in the ganzfeld.
- Elementary changes of sensory qualities are usually observed
already after a relatively short exposure to the visual or MMGF (a few minutes). The visual field’s luminance diminishes and the field shows diffuse inhomogeneities, often described as a ‘cloudy fog’. In case of a colour ganzfeld, the field’s colour gradually bleaches, up to the point of a loss of the sensation of colour: the field is of indefinite grey, sometimes with an undertone of the complementary colour, e.g., greyish-green if red light is used. In addition, more distinct structures may appear against the diffuse ‘foggy’ background: dots, zig-zag lines, or more complex patterns. Generally, these elementary perceptual phenomena can be accounted for by adaptive retinal processes: saturation of the receptive elements and their mutually inhibitory interactions
- After a prolonged exposure (a few minutes up to tens of
minutes) to the ganzfeld, some subjects report complex percepts
- Another phenomenon occasionally reported from the
ganzfeld are episodes of ‘‘complete disappearance of the sense of vision for short periods of time’’, also called ‘blankouts’ (Cohen, 1960), occurring after prolonged exposure (10– 20 min) to the ganzfeld. Subjects also report that during these periods they were uncertain whether their eyes were open or closed, or even unable to control their eye movements. In the ‘luminous fog’ of the ganzfeld the subjects do not see anything; in the ‘blank-out’ periods, they may experience presence of ‘nothingness’ (Gibson, 1979).
- Interestingly, the very first hallucinatory percept of the
three reported above emerged after a ‘blank-out’ period. Herrmann (2001) in an electroencephalographic (EEG) study of the visual cortex’s response to a flickering visual field observed the appearance of subjective colours and forms. Herrmann and Elliott (2001) described the variety of these perceptual phenomena as a function of flicker frequency (1– 40 Hz). Recently, Becker and Elliott (2006) reported cooccurrences of forms and colours in a flickering ganzfeld being dependent on flicker frequency, and phase relationship between the subject’s response and the flicker period.
- Immobilised retinal images
The image created by the eye’s optical system can be fixed on the retina by special techniques (Heckenmueller, 1965). The structure of the visual field thus remains preserved but the scanning motion due to eye movements is inhibited. Under these conditions, partial or total ‘fade-outs’ of the visual field may occur (Yarbus, 1967), indicating that a regular refreshing is necessary for maintaining the visual structure. We may hypothesise a relationship between these ‘fade-outs’ and the ‘blank-out’ periods in ganzfeld, where eye movements are reportedly reduced.
- However, later studies
revealed functional differences between sub-bands within the alpha frequency range: low-frequency alpha, reflecting rather attentional processes, and high-frequency alpha reflecting cognitive processes (Klimesch, 1997, 1999; cf. also Shaw, 2003).
- Cohen and Cadwallader (1958) reported correlation between
higher alpha activity in the resting EEG and individual susceptibility to ‘blank-outs’. Cohen (1960) interpreted occurrence of alpha activity during the ‘blank-outs’ as alpha rebounddthis is a well-known phenomenon where, after a transitory suppression e.g., due to an external stimulus, eyes opening, etc., alpha activity attains the original level, or even increases. Tepas (1962) found an increase of alpha amplitude during the ‘blank-outs’, which was intermediate to ‘eyes closed’ and ‘eyes open’ conditions, but could not confirm the hypothesised relation between high alpha activity and blank-out susceptibility. These findings are in line with early observations by Adrian and Matthews (1934), who had previously reported alpha rebound after eyes opening in a uniform visual field. Later, Lehtonen and Lehtinen (1972) also reported re-occurrence of alpha activity in the ganzfeld, comparable to the ‘eyes closed’ condition. Increase of alpha activity was also observed during the ‘fade-out’ periods in perception of stabilised retinal images (Lehmann et al., 1967); this supports the relation to ganzfeld ‘blank-outs’ hypothesised above. As shown in the preceding sections, the variety of ganzfeld- induced phenomena is fairly rich and suggests relations to several different classes of perceptual phenomena and/or states of consciousness. Objective characterisation of the brain’s functional states under ganzfeld stimulation by means of EEG measures may help to elucidate these relations. This was the objective of our two major ganzfeld studies, results of which are summarised below.
- There is recent experimental evidence
that eyes-open and eyes-closed conditions are not equivalent even in the absence of any visual input (Marx et al., 2003).
- Within the waking states, the ganzfeld and ‘normal’
waking states were best distinguished by the band power ratio a2/a1 (frequency ranges 10–12 Hz and 8–10 Hz, respectively), which was increased in the ganzfeld EEG, indicating an acceleration of the alpha activity. Visual inspection of the spectra reveals a power drop along the lower flank of the alpha peak in the ganzfeld EEG, leading to an increase of the peak frequency
- to ganzfeld imagery, with the maximum alpha acceleration in
the time segment 20–10 sec before the report. A time–frequency analysis of the data gave additional evidence for alpha acceleration (Wackermann et al., 2003).
- GFI–EEG spectra and revealed various forms of ‘correlation
profiles’ over the analysed 30 sec time window. The most stable correlation over this time window was a global (i.e., involving all 19 channels) negative correlation between a2 power, measured relative to individual GFB baselines, and subject-reported vividness of imagery.
- The relation between fast a2 activity and imagery formation
was interpreted by Pu¨ tz et al. (2006) as an indicator of activation of thalamo-cortical feedback loops involved in retrieval, activation and embedding of memory content in the ganzfeld-induced imagery. The observed a1 attenuation during the analysis epoch may reflect a shift of attention towards the visual percept and, later, preparation of the required motor action (button press signalling occurrence of imagery). The unspecific alpha-inducing effect of the ganzfeld-induced steady-state (no imagery) is in line with the inhibition hypothesis (i.e., alpha synchronisation due to inhibition of cortical areas related to external sensory information processing), and with earlier findings of other authors mentioned above
*comparing electrophysiological signatures of sleep onset, waking mentation and ganzfeld, it was established that ganzfeld imagery occurs in a brain functional state which is distinctly different from that occurring at sleep onset
* The subjects’ eyes were covered by translucent, anatomically shaped goggles, which were illuminated by bright red light. Simultaneously they were listening to the monotonous sound of a waterfall through headphones. Sound intensity was individually adjusted to a subjectively comfortable level. This procedure has proven to accomplish an almost perfect homogenisation of visual and acoustic sensory field
*alfa1/alfa2 location Pz *a1 power shows a minor but continuous decrease during the entire 30-s analysis interval,while a2 displays a distinct tri-phasic time course: initially (30 to 21 s) reduced power, relative to baseline level; followed by a F burst_up to the baseline level in the second sub-interval (20 to11 s), and finally a monotonic and strong decrease during thelast sub-interval (10 to 2 s). These time courses are reflected by the corresponding Z values *differences in the time course of the two a sub-bands are particularly well expressed by the a2/a1 ratio , 8–10 Hz (a1), 10–12 Hz (a2) (a2/a1=-2)
*negative coreleation of a2 power *There was a positive global correlation for variables b3 power and vividness of imagery
Gamma /theta synchrony
Tetris effect - Cognitive overload
Hypocampal buffer
MSI
Neuromodulation
- cholinergic
- dopaminergic
Subjective measuring
Siganl priority - P
- Preception priority of signal 0-5 , 3 is border of consciousnes preception .
Signal Intensity - I
- 0-100
Use of system resources - R
- 1R = 1/100 of resources aviable in normal state of wakefullnes
Image properties
- Phase

- Brightnes

- Size % of visual field

- Time
- 0,5 flash - image unrocgnizable
- 1 flash - image rocognizible
- 2 image
- 2,5 series of image
- 3 scene
- 4 micro dream
- 5 dream
- Theme stability
Objective measuring
?
Expediments
- mainly subjective data from automatic drawing , greaphs drawn in PC was simplified
Eye blink effect
- First i notice this in teris effect tests .When i blink i notice very short “flash” of image .First idea was that is becouse closed eyes but when i closed eyes for longer time i not see any new images or see only much weaker images than on blink (depends os strenght of tetris effect).
Change of non-physical vision datastream priority when blink - in normal state
- base priority NV around 3,83
- duration 20-150ms depend of blink time
- NV in peak 3,1 - 3,2
- after blink 0,01 - 0,03 lower than base

- time in s , priority
Change of non-physical and physical vision datastream priority when blink - in normal state
- + try to get beter temporal resolution

- time in s , priority ,green physical vision
Change of non-physical vision datastream signal intensity when blink - in normal state
- normal intensity 0,058
- longer blink higher intensity increase by similar rate
- back to normal 12-15ms
- TO DO : longer eye closed , synchornization with priority changes

Change of non-physical vision datastream signal intensity and priority when blink - in normal state

- blue line eye closed/open
- change of priority not depend on duration of blink
Change of non-physical vision datastream used system resources and priority when blink - in normal state

Slow eyes closing and opening
- test various closing and opening rates

- one “box” = 0,1s
- priority of NV
- why NV priority decerase when eyes close, what presisly cause this effect ? Slower closing better and priority? + TEST even longer closing ,P-FV,R-FV,R-NV,I-FV,I-NV

- reason find : decrease is caused by focus on FV behaind eylids blacknes
- no focus on NV because no precevable NV , increase NV by disinhibition ?
Pasive relaxation

- N- nonpysical
- F- physical
- T- touch+termo
- P- proprioreception
- V- vision
- in end strong hypnagogic halucination but low lucidity
Pasive vs progresive relaxation - test on hands
- deapnes of hand relaxation was very similar but interesting is second images witch show changes in preceprion priority
- senory intereaction and turning on and off of senses, many intresting things to research
- 3 is a limit of consciousnes preception when signl break this barier down to higher priority is consciously precieve, when signal move above 3 is not longer precived
this chnage is noticable in relaxation but is hard to describe it its like change from 2D to 3D balacknes diffent preception of space .This change have different levels i suspect it depends how much senses is turn off (vision , proprioreception..)


Pasive relaxation - red behaind the eylids
- intreaction of thoughts , NV and FV priorities

Pasive relaxation - change of position , hypnagogic state, sleeap signals , dream
- about 30 mins of pasive relaxation before recording “image” , i try to sleeap and change position minimal 3 times (left,right,left)
- i notice some hypnagogic imaginery before recording
- recording start few second before chnge position to laing on left side (red dots - eyes open)
- brown time line is changing position forehed against wall of the bed - notice blue line - physical touch in priority
- greean line -physical vision
- light blue time line - stable hypnagogic satate constatnt flow of images and short scenes ,hyp phase about 0,5-1,5 (depend of NV priority),orange line in P
- yelow time line + blue line - Sleep signals ,touch based dream , somthing else? This need more data
- Blakc time line in R - Probalby lost of consciousnes or very weak
- Indigo time line - dream , notice also increase in R
- Unfortunetly i recording only NV,FV,FT and R and large part of intresting data is lost, experinc continue about next 20min dream,hypnagogic stability control , 2x false awakening .Without data from other senses is FV,NV almoust meaningless

Visualisation : Visual attention focus problem

31.5 update
EOG and automatic drawing test 10min
- Automatic drawing of FV and NV
- horizontal EOG
Goal : compare EOG (delta amplitude ) to increase FV priority
Result :
- all FV priority peaks found in EOG (+-5s drawing is not that precize ,recording about 9min )
- few peaks in EOG but not in drawing –) only in pars where priority is high and eye movemt or focus to FV no longer incerease it OK
Audio EOG biofeedback to minimalize eye movements
Pasive realx + sugestion not to move eyes and breathing to eyes about 1 min

- preception priority P (0-5, lower have higher priority )
- NV non-physical vision
- FV physical visoon
- P-FV up -) P-NV down
- P-NV up -) P-FV down
- P-NV up peaks (value decrease) = “hypnagogic images”
- P-NV large down peaks or P-FV large up pekas = eye movement of focus to FV
- orange arrows area = behaind the eyelids backnes in this cases rednes (focus to FV)
- orange points P-FV up and P-NV up = other sense influence,thoughts,integration .. (need more research)
Pasive realx - morning

- red dot = sneeze
- pimk dots = large eye movements (also possibe focus to FV without the movement but less probable )
Active realx - focus to breath and heart beat

- evening 20:30
- hard to focus
- chaotic thoughts
- cognitive overload (tetris effect)about 6h watching new TV serial , this increase base P-NV
- P-NV higher than 3 after 15s (probably due the cognitive overload)
- T 2,75 P-NV and P-FV decrease probaly due focus to other senses or thought
- T pink line = auto eyes open , few times durring the session my eyes slightly open by themselves
- T green line = dream scene , no loger flashes of image but scene (may be caused by visualization , automatic asociation about thought tema , or themselfs)
visualized ones have higer P than asociation , asociation higher than automatic (in similar time in one session )
- P-FV below 3 = very important moment mean physical vision is turn off and allow NV disinhibition
Active relax - focus to breath and heart beat - test 2

- same day evening 22.00
- more awake than in previous test but less images
- P-NV higher than 3 after about 18s (probably due the cognitive overload)
- T 0:18-0:45 asociation created images i remeber few names from serial
- T 1:08 P-FV and P-FV go down probably because start focusing to breath and heart rate (probably increase of propriorecoption P )
- T 4:45 P-FV uder 3
- T 6:22 - 9:00 probably proprioreception effect
- gereen line above T 12:30 active vizualization of flight on dragon (scene form serial) ,2 scenes before caused by mix of visualization nad asociation (more asociation)
Some test (second part of larger test unfinished )
- before drawing 12 min of relax and eye open

- T around 1:45 green line = vizualization -) images
- T around 2:00 yellow line = after vizualization automatic asociation nad scene with lower P
Realx - Breath - morning
- 9:30h

- shorter inter flash intreval than in other test (morning ?)
- P-NV stabil larger than 3 1:40
- P-FV below 3 : 5:00
- T red line = behaind the eyelids blacknes in this cause redness in this sessoin cause problems (too much light in the room)
- T 10-11 focus to FV redness
- viz = active vizualization
EOG biofeedback
- audio EOG SD1s amplitude(4,5uV+) to volume as feedback

Realx - guided 7min
- guided relaxation

- FP physical proprioreception
- P-FP allow distiguish between eye movement and focus without movement
- T 10:45 - 11:30 increase of P-FP : focus to body ? cause P-FV go below 3 ? –) need more tests
TO DO –)increase vertical resolution and add more senses
2.6
notes:
- Fleeting vs Fully-Formed Hypnagogic Images.
'fleeting' vs. 'fully-formed' images. These two image types are distinguishable by several qualities, including the following.
- 1)
Temporal. Fleeting images tend to occur during the first stages of a self-observa tion trial, before a fully-formed image has arisen.
- 2)Duration.Fleeting images are brief; they appear and disappear in an instant, often they are not detectible during
self-observation without a firmly established intent. Fully-formed images are more enduring and thus more easily observed and recalled.
- 3)Simplicity
. Unlike a fully-formed hypnagogic image, a fleeting image may occur in only a single sensory modality. It may consist of nothing more than an isolated flash of visual scenery, a single fragment of spoken voice, or a brief moment of kinesthetic sensation. Fully-formed images may be comprised of all three modalities.
- 4)
Ineffability . Because they are brief, simple and outside the range of most observers' waking and dreaming experiences, fleeting images may easily defy verbal description. Momentary kinesthetic sensations are particularly difficult to describe because there is a lack of a commonly accepted vocabulary for the domain of kinesthetic experience. Fully-formed images more nearly resemble nocturnal dreams and are similarly more amenable to description. To characterize the nature of fleeting images adequately, an observer must often advance either metaphoric discourse or completely novel terminology.
- 5)
Association with sleep onset feelings. As shown in Table 2, fleeting images are often completely inseparable from subtle sleepiness feelings, as if the images were an integral part of sleepiness itself. Although sleep onset feelings may also accompany fully-formed images, in these cases they are often not obvious. Their ephemeral nature may be masked by the dramatic complexity of more fully-formed multimodal imagery
- Phasic neuromuscular events (NMEs) which regul
arly accompany hypnagogic images, such as limb jerks, were found to be isomorphic with some imagery content. In Study 1 (fully-formed images), phasic NMEs were found to accompany 56.7% of images. In Study 2 (fleeting images), this proportion was only 16.7%. However, the proportions of phasic NME types for the two classes of imagery were strikingly similar: head tilts (61.5% v. 56.4%), leg jerks (23.1% v. 23.1%), arm jerks (7.7% v. 15.4%) and whole body jerks (7.7% v. 5.1%) respectively. It seems likely that many NMEs accompanying fleeting images were too subtle or transient to be detected by this self-observation technique, a possibility supported by physiological recordings, such as those in Study 3.
- Although the duration of the pre-image interval was
variable, the pattern of EEG changes was similar from image to image. These findings suggest that the production of hypnagogic imagery for this observer may have a more or less invariant underlying physiological pattern. The relationship of this pattern to that of the previously-described phenomenological events remains tobe clarified. However, it seems likely that the observed in EEG activity correspond to periods of activation and deactivation of hypnagogic imagery processes * In most cases, cessation of eye activity was followed by a diminution in alpha activity and EEG amplitude across most channels and an increase in the prevalence of other frequency bands.


4.6
Lucid Dream by use of multisensory integration NV scene H 2-3 stabilized by sense of touch in legs to H5. Some route in forest near water i look at feet and focus to sense of touch (to grass , groud)it takes bout 20s to stabilize to H5.LD have about 5 parts whn drem destabilize i stabilze it again sometimes i almoust lost sense of vison. * i have EEG recording form Fp1 and T5 somwhere in it is LD recorded
Recording:
- CH1 Fp1 A1
- CH2 T5 A1
- GND A2


- relaxation on back
- 40 move head to right side (stil on back)
- 50 move to side
- 65 big change (62:40-65:00)
- 65-75 start of LD (not enought data and experinece to determine if start 65 or 75min) ,(automatic drawing show strat 65 but information show only NV without inromation about R and other importatnt to detemine LD)
Recording for Automatic drawing was turn OF i try reconstrut some inromation from memory but resutl gave many gaps and i doun tknou how reaiable it is .
Sleep spindle
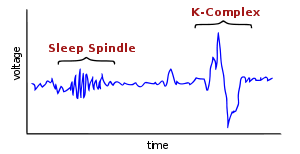
- Sleep spindles (sometimes referred to as “sigma bands” or “sigma waves”) may represent periods where the brain is inhibiting processing to keep the sleeper in a tranquil state. Along with K-complexes they are defining characteristics of, and indicate the onset of, stage 2 sleep. They are often tapered at both ends and frequently seen over the frontal and central head regions. They may or may not be synchronous, but they should be symmetrical and bilateral.
- During sleep these spindles are seen in the brain as a burst of activity immediately following muscle twitching.
- The greatest spindle activity occurs at the beginning and the end of the non-REM portion of the sleep cycle.
- There are also slow spindles in the range of 11 - 13 Hz that are also associated with increased activity in the superior frontal gyrus, and fast spindles in the range of 13 - 15 Hz that are associated with recruitment of sensorimotor processing cortical regions, as well as recruitment of the mesial frontal cortex and hippocampus. What do these spindles mean? Ongoing research hopes to illuminate their function.
- Recent research by Matthew Walker and his research team at the University of California Berkley shows that sleep spindles are associated with refreshment of our ability to learn. This research showed that the greater the number of sleep spindles produced by napping participants, the more they were refreshed to perform on a learning task. Further, the brain areas most involved were the hippocampus and the prefrontal cortex. These are areas that are critical for learning.
- The incidence of spindles is reported to peak at a frequency of 0.2-0.3 Hz
- The incidence of spindles has considerable variance (1±40 s inter-spindle intervals in humans). Also variable is their topographic prevalence in the brain, their time of appearance in sleep stages, their association to other EEG landmarks (like K-complexes) and their dependence on drugs. All these suggest that spindles do not constitute a unique and/or uniform phenomenon.
- The association of spindles the slow cortical oscillation (~0.75 Hz) [12] is proposed to be causative in the sense that this oscillation which is supposedly generated within neocortical networks, synchronizes neuronal activity into generalized down-states (hyperpolarization) of global neuronal silence and subsequent up-states (depolarization) of increased wake-like neuronal firing. With the beginning of the latter cortico-thalamic volleys are proposed to drive the generation of spindle activity.
- Such a strong and synchronous input may discharge reticular cells directly or indirectly and thus could serve as the initiator of sleep spindles [2]). In a recent study the incidence of spindles immediately following K-complexes was between 65-70%
- However in this study neither the probability of appearance nor the power of spindles correlated to the amplitude or any other feature of the K-complexes that preceded them. When K-complexes appeared spontaneously after the start of a sporadic spindle, the spindles were invariably shut down for the duration of the K-complex, usually being replaced by a short lived oscillation in the high theta frequency band. Also the spindles appearing immediately after a K-complex had invariably faster spectral frequency than the sporadic spindles. Such findings suggest that the association of K-complexes with spindles is strong but may be due to a common trigger rather than a causative interaction.
- Animal and human studies show that spindles are sleep maintaining events [65-66] that block the transfer of sensory information to the cerebral cortex during sleep [64], thus preventing sleep-interrupting arousals.
- While TC neurons are hyperpolarized and engaged in this bursting mode, sensory afferents are expectedly prevented from reaching the cortex, resulting in almost complete deafferentation, except for very strong or alarming stimuli [65].
- The recent observation (described in 2.2.) that spindles are invariably shut down for the duration of the K-complex and they appear right after at increased spectral frequency [71] support a role of spindles in preventing stimuli (which triggered the K-complex) to reach consciousness.
- Anesthetics lead also to loss of consciousness - in the sense of turning the subjects oblivious to their environment - with different mechanisms depending on the drug used. One mechanism they partly share with natural sleep is apparently the production of spindles in a similar way, as several anesthetics hyperpolarize TC neurons (by activating 2PK channels and/or by potentiating GABA receptors) and halothane induced spindles are antagonized when carbachol is injected into the pontine reticular formation [74]. Spindling then causes a decorrelation between sensory input to TC neurons and these neurons’ output to the cortex, thus contributing to the loss of consciousness.
K complexes
- are large waves that stand out from the background and often occur in response to environmental stimuli such as sounds in the bedroom.